
|
|
|
Sierra Nature Notes, Volume 3, May 2003
How
has climatic variation influenced treeline dynamics in the past?
Andrea H. Lloyd
Assistant Professor, Department of Biology, Middlebury College
Middlebury, VT 05443
We reconstructed a 3500-yr history of fluctuations in treeline elevation and tree abundance in the southern Sierra Nevada. Treeline elevation was higher than at present throughout most of the last 3500 years. We studied foxtail pine (Pinus balfouriana) stands at the crest of the Sierra Nevada, where warm temperatures (ca. 100 BC to AD 900) allowed trees to migrate upslope above current treeline (the upper-most limit in elevation that upright trees exist). Subsequently, periods of severe droughts (AD 1000 to 1400) or cool temperatures (AD 1500 to 1900) caused treeline to move downslope to its current location.
 |
| Foxtail Pine (Pinus balfouriana) north of Mt. Whitney, Sequoia National Park. Tree ring cores of these trees — living and dead — extend our knowledge of Sierra climate back thousands of years. |
How do we study treeline dynamics?
In the cool, dry climate of the Sierra Nevada, dead foxtail pine
trees at high elevations are preserved in situ for millennia. Dead trees
located
above current treeline testify to the dynamic behavior of treeline in
the past. In order to date each tree, we extracted increment cores from
each
tree and used tree-ring
dating techniques to identify the calendar year
of each tree's inner and outer ring. In order to document general trends
within Sequoia National Park, we sampled five sites (Table)
on the eastern crest of the Sierra Nevada (see Map).
Our ability to interpret treeline
movements
in terms of past climatic variation was made possible by the results
of SNGC studies of past climatic variation.
By comparing estimates of the numbers and positions of trees above and at treeline under different climatic regimes in the past, we can infer how climatic variation affects treeline dynamics. We see two distinctly different processes at work in our study areas. During some periods in the past, severe drought has reduced treeline populations by causing the death of adult trees. During other periods, low temperatures reduced the number of trees at treeline by inhibiting recruitment of seedlings. However, because foxtail pine can live for more than 1000 years, short-term failures in recruitment rarely causes large changes in the structure of these treeline forests.
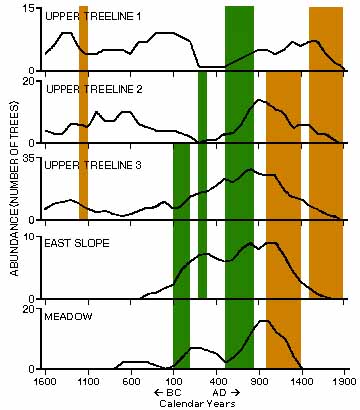 |
| Abundance
of live trees in study plots in currently unforested areas. Abundance
is the minimum number of trees that were alive
during any part of each 100-yr time interval. Dates on the x axis are
the first year, in units of years BP, of each 100-yr time interval; A.D.
1950 is the zero year. Thus 50 yr BP is the interval from AD 1900 to
present. Note that the scale of the y axis differs among sites. The green
bars are time periods when there are significant increases in tree
density (i.e., treeline advanced) and the brown bars are significant
decreases (treeline declined). Figure taken from Lloyd and Graumlich. 1997. Holocene Dynamics of Treeline Forests in the Sierra Nevada. Ecology 78(4): 1204. |
What are the implications of this study for understanding the future of
the Sierra Nevada?
Seedlings are currently being established above treeline. From our studies,
we know that this incipient treeline movement reflects the relatively warm
and moist conditions that have persisted for much of the 20th century.
And, we know that these changes are still within the natural variability
we have
witnessed at these sites for the past 3500 years. Our results
have three primary implications for the potential of anthrpogenically
induced warming to cause future changes at treeline.
First, although large changes in forest structure and distribution occur on time scales of hundred of years, the rate at which these forests respond to rapid future climate change is likely to be slow, and treeline position may lag climate change by decades to centuries. Monitoring efforts at treeline might therefore emphasize change in density of existing stands as well as change in position of treeline.
Second, the period from AD 1050 to 1450 illustrates the extent to which water balance can reverse treeline response to temperature. Whereas climate appears to have been warm when treeline forests expanded, warmth does not necessarily lead to subalpine forest expansion. The observational record of the relatively wet 20th century leads to the prediction that treeline will rise if temperatures rise. The paleoecological record, however, indicates that future warming is unlikely to cause an expansion of subalpine forests if it is accompanied by a reduction in water supply.
Third, responses to past climate variation have involved more than simple upward or downward movements of the treeline boundary: they have involved relatively complex changes in distribution, accompanied by changes in forest structure. The impacts of even minor changes in the elevation of treeline are therefore likely to be substantial. A warming-induced rise in treeline elevation is likely to involve landscape-scale increases in biomass, productivity, and carbon pools as a result of increases in both forested area and density of current forests.
Editor's Note: This article first appeared, in a slightly different form, on the USGS Sierra Nevada Global Change Research Web Site.
Acknowledgements
For field assistance we acknowledge Alison Arians, Kirsten Prettyman, John
King, Paul Sheppard, Carol Pranka, Lisa Graumlich, John Lloyd, Erin Lloyd,
Peter Lloyd, Chris Fastie, Annie Esperanza, and Nate Stephenson.
For Further Reading
Lloyd, A.H. 1997. Response of treeline populations of foxtail pine (Pinus
balfouriana) to climate variation over the last 1,000 years. Canadian Journal
of Forest Research 27: 936-942.
Lloyd, A.H. and L.J. Graumlich. 1997. Holocene dynamics of treeline forests in the Sierra Nevada. Ecology 78: 1199-1210.
Graumlich, L.J. and A.H. Lloyd. 1996. Dendroclimatic, ecological and geomorphological evidence for long-term climatic change in the Sierra Nevada, U.S.A. in Tree-Rings, Humanity, and the Environment: Proceedings of the International Tree-Ring Conference. Tucson, AZ. 1994.
Sierra Nature Notes:
Sierran Treeline Dynamics in a Changing Climate by Andrew G. Bunn
The Great Droughts of Y1K by
Scott Stine, Ph.D
Maps & Tables
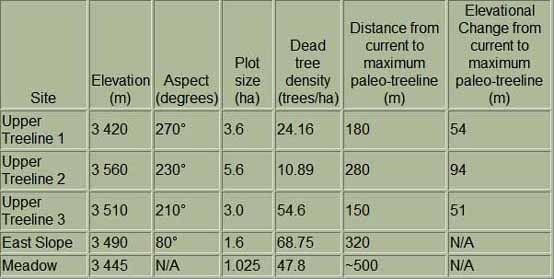 |
Study Plots |
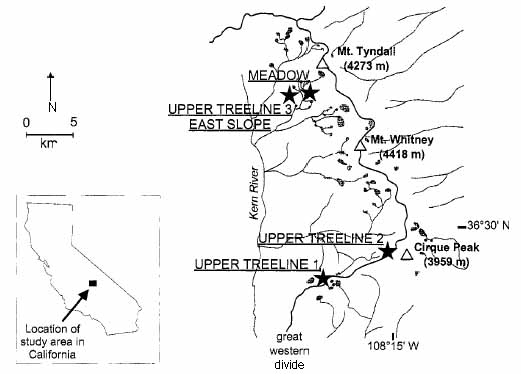 |
| Location
of study sites in Sequoia National Park. The inset shows the location
of the study area in California.
River drainages are indicated by the light gray lines. The Great Western
Divide, indicating the major crest of the Sierra
Nevada, is indicated by the solid black line. Major peaks in the area
are indicated by triangles. Stars show the position of
sites. Upper Treeline 3 and the East Slope sites are <1 km apart,
so are indicated by a single symbol. Figure taken from Lloyd and Graumlich. 1997. Holocene Dynamics of Treeline Forests in the Sierra Nevada. Ecology 78(4): 1201. (Back to text) |
Our Founder Questions? Go to About Our New Site |
Masthead
Photo from: |
